
木质纤维素因存在复杂的多尺度抗降解屏障,其酶解糖化效率受到限制。热水预处理(HTP)作为主流的预处理技术,虽能显著提升糖化率,但在机理研究方面仍面临诸多痛点:
1、湿态结构表征难:传统 BET、MIP(汞压法)在测试前需干燥样品,这会导致孔隙坍塌,无法真实反映酶解过程中生物质的“水合可及性”。
2、单一尺度局限:SEM 仅能观察样品表面形貌,XRD 仅能测定结晶度,难以对细胞壁内部 5 – 50nm(酶分子尺度)的有效通道进行量化分析。
3、可及性量化不精准:无法有效区分哪些孔隙是酶能够进入的活性位点。
4、测试周期与代表性:传统溶质排斥法(SRA)操作繁琐,且无法原位监测水分动力学变化。
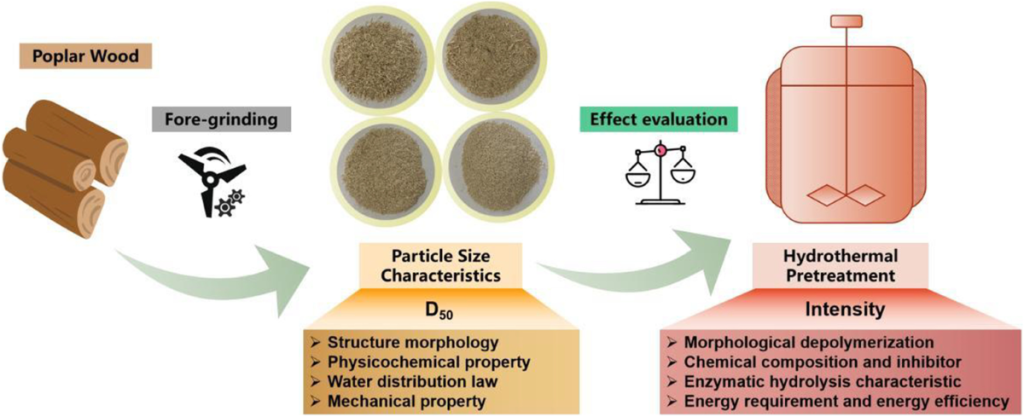
图 1 (Graphical Abstract / 流程示意图)
本研究采用纽迈分析(NIUMAG)生产的低场核磁(TD – NMR)设备(如 VTMR20 系列),通过T2弛豫分析与核磁孔径分析(NMR Cryoporometry, NMRC)联用的方式,在不破坏生物质天然水合状态的前提下,构建了“预处理 – 微观结构 – 糖化性能”的定量逻辑链:
水分动力学分布:可区分结合水、细胞壁孔隙水、自由水。
纳米级孔径分布曲线:实现 5nm – 100nm 的连续量化。
纤维素有效可及表面积(ASA):准确评估纤维素的可及程度。
多尺度结构综合评估:结合XRD测定结晶度,全面了解生物质结构。
将热水预处理木材的水分分布划分为三类:
结合水(T21):紧密束缚于细胞壁聚合物。
孔隙水(T22):位于细胞壁内新形成的微孔中。
大孔/间隙水(T23):位于管胞腔或颗粒间隙。
通过纽迈 TD – NMR 的 CPMG 序列测试,研究发现热水预处理过程会直接重塑木材的水分分配模式:
1、低运动性相(T21结合水):
随着热水预处理强度增加,T21峰位左移(弛豫时间缩短)。这表明半纤维素脱除后,剩余纤维素束之间的氢键作用增强,导致水分受限程度更高。
2、关键中间相(T22细胞壁孔隙水):
这是评价糖化效率的核心指标。实验数据显示,热水预处理后A22(峰面积)显著上升,且T22弛豫时间从几毫秒扩展至数十毫秒。这证明热水预处理产生的“体积膨胀效应”在细胞壁内创造了大量纳米级孔隙,显著增加了水分的流动空间。
3、高运动性相(T23自由水):
A23的增加反映了热水预处理对木材宏观结构的粉碎程度,颗粒粒径减小,暴露出更多的外表面积。
技术价值:T22组分的演变直接量化了细胞壁的“开启程度”,为预处理工艺的筛选提供了直观的物理判据。
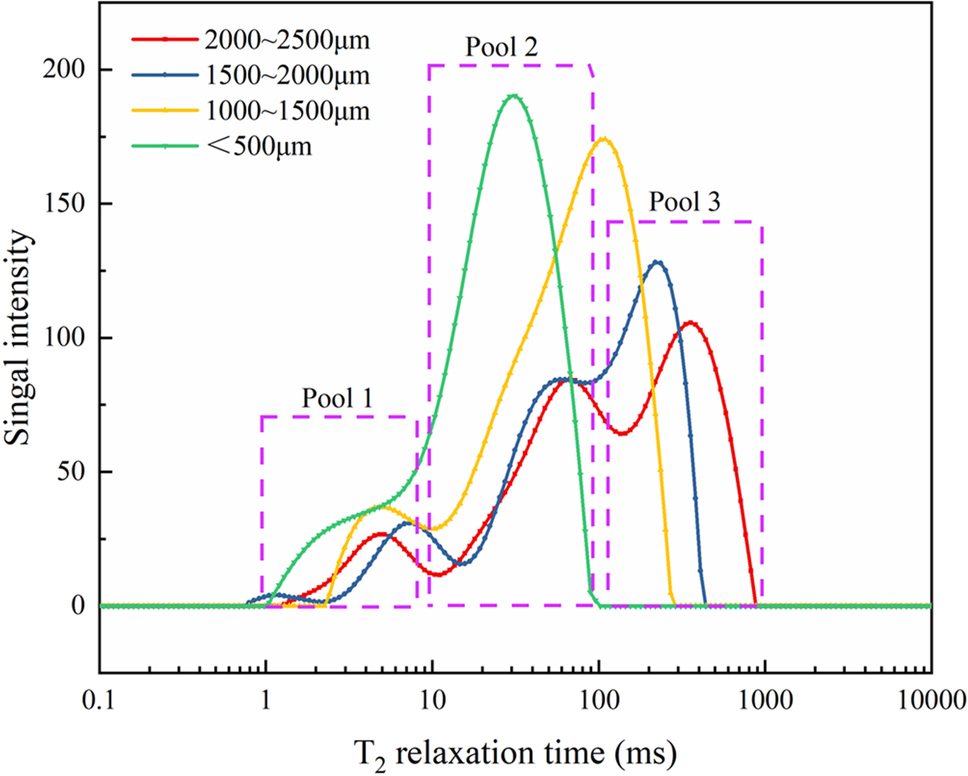
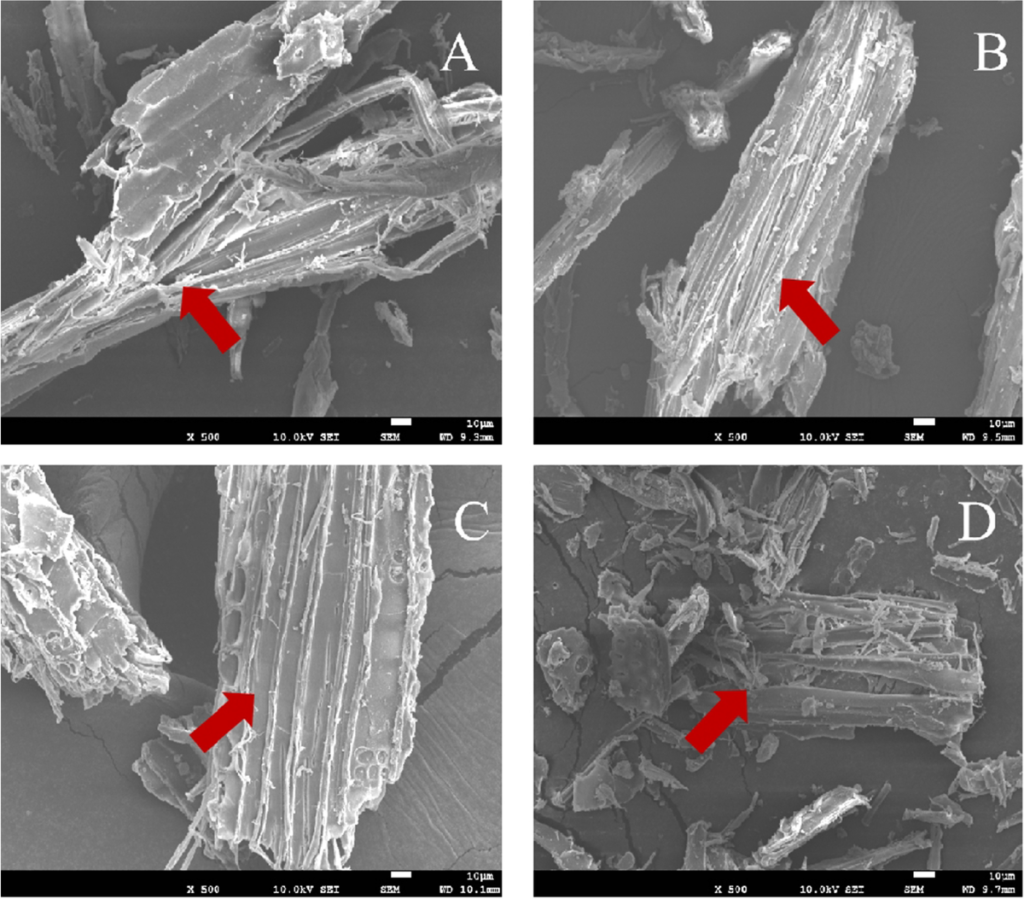
图2 (T2弛豫谱图) 与 图 3 (SEM 图像)
利用纽迈设备的 NMR Cryoporometry 技术,通过监测孔隙内水分的相变(融化过程),实现了对湿态木材孔径分布的精准表征:
1、孔径分布(PSD)位移:
对照组(天然木材)孔径主要分布在 < 5nm。热水预处理后,5nm – 50nm 范围内的孔体积显著增加。由于纤维素酶的流体力学直径约为 5.1nm,这一区间的孔隙才是酶解的“有效通道”。
2、有效可及性量化:
通过 NMR 曲线积分计算出的有效孔体积与最终的酶解产糖量呈现高度线性相关(R2>0.95)。
3、多尺度互补:
结合XRD(X射线衍射)发现,尽管热水预处理可能提高纤维素结晶度(通常不利于酶解),但 LF – NMR 测得的可及性大幅度增加弥补了这一缺陷。这说明:对于热水预处理木材,孔隙可及性是主导糖化效率的首要因素,而非结晶度。
技术价值:NMRC 填补了传统表征无法测量“湿态有效孔径”的空白,为解析酶解屏障提供了关键证据。
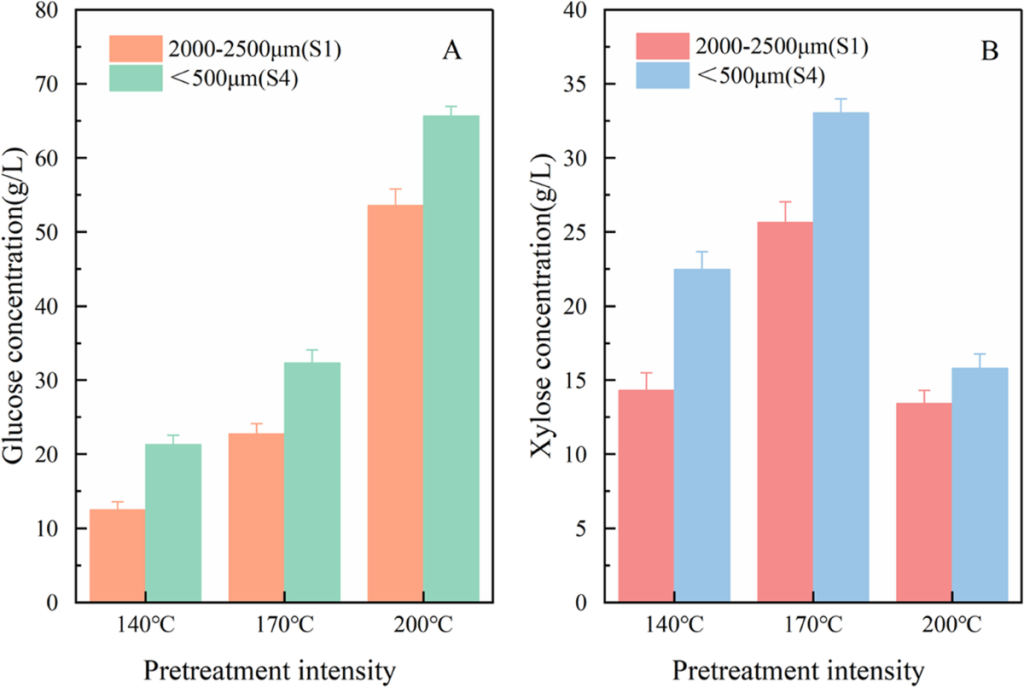
图4 – Glucose Yield
1、实现微观指标与生化产率的预测性关联
研究证明,纽迈 LF – NMR 测得的T2谱图总面积及T22峰值可直接作为预测糖化效率的“指纹指标”。通过这些参数,可以在进行长达 72 小时的酶解实验前,仅用 2 分钟测试便预判预处理效果,极大缩短了研发周期。
2、建立真实环境下的原位表征体系
该技术体系解决了生物质研究中“干湿态不一致”的难题。低场核磁测试无需复杂制样,重复性好,日间偏差小,实现了从“静态形貌观察”向“动态动力学量化”的技术跨越。
3、构建多尺度结构的完整画像
纽迈低场核磁与高场核磁的“高低结合”,能够同时提供分子尺度(结晶度、化学键)与超分子尺度(孔隙分布、水合可及性)的信息,为木材科学研究提供了全尺度的解决方案。
《Evaluation of initial material particle size on the hydrothermal pretreatment of poplar powder》
 扫描二维码
扫描二维码 扫描二维码
扫描二维码电话:400-060-3233
售后:400-060-3233


返回顶部